


This article is a primer on water vapor transpiration in plants, one of the most important phenomena in the hydrologic cycle. We begin with a brief discussion concerning stomata, the adjustable pores that mediate gas exchange between leaves and the atmosphere. A quick overview of the stomatal opening mechanism and the factors that affect it is provided. The resistance/conductance of stomata and other components of the gas-exchange pathway are presented, and the text closes with a simple set of calculations involving these parameters.
1. Stomatal anatomy and aperture
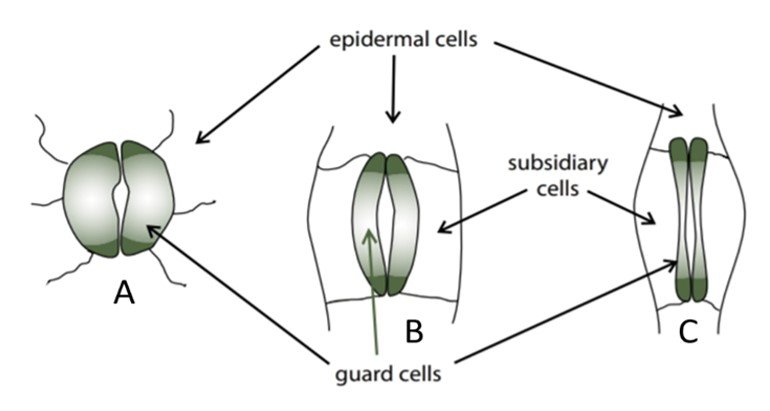
Stomatal anatomy varies from one plant to the next, and more primitive species often have less refined stomata. Figure 1, extracted from Willey’s Environmental Plant Physiology, illustrates the stomata of three types of plants. Illustration A shows the stomata of primitive plants, which have no dedicated subsidiary cells and produce a relatively small cavity even when guard cells are turgid. Illustration B shows the stomata of many angiosperms, in which case the guard cells are accompanied by subsidiary cells that work together to produce a larger cavity. Illustration C shows the stomata of the grass Triticum aestivum, whose dumbbell-shaped guard cells operate in tandem with the subsidiary cells to open along their length, maximizing the area of the stoma. Grasses have characteristic guard cells and rapidly responding accessory cells, factors that partially explain their high water use efficiency (WUE).
Figure 1. Anatomy of stomata.
At full opening, the stomatal apertures of Phaseolus vulgaris (bean plant) measure 3

The stomatal aperture is controlled by the conformation of the two guard cells surrounding a pore. These cells are generally kidney-shaped, may be 40
Figure 2. Opening and closure of stomata. Effects in blue chronologically occur from top to bottom.

It was long assumed that stomata respond homogeneously over the entire leaf. However, leaves of water-stressed plants exposed to 14CO2 often show a heterogeneous distribution of fixed 14C. This shows that some stomata close completely (there is no radioactivity close to these stomata), whereas others hardly change their aperture (radioactive label is located near these stomata). This patchiness of stomatal closure can also be visualized dynamically and nondestructively with thermal and chlorophyll fluorescence imaging techniques.
2. Factors that affect stomatal opening
Several factors affect stomatal opening or closure to varying degrees. Some are briefly discussed below.

Atmospheric humidity: In addition to assimilating carbon dioxide, stomata also mediate transpiration of water vapor to the atmosphere. The water vapor content within the leaf is usually close to saturation. So, a dry atmosphere gives rise to a large gradient in water vapor across the stomatal pore, that is, a leaf-to-air vapor pressure deficit (VPD), which increases as the relative humidity outside falls. Increasing VPD leads to a proportional increase in transpiration rate through the stomatal pore, driven by diffusion. Stomata therefore generally close at high VPD to prevent excessive water loss.
Carbon dioxide: Stomata react to the gradient of CO2 concentration between the external air and the intercellular spaces of leaves. Decreasing CO2 concentration in ambient air causes stomata to open, while increasing it causes stomata to close. The degree of stomatal opening often depends on the concentration of CO2 in the guard cells, which reflects their own carbohydrate metabolism as well as the CO2 level in the air within the leaf. For instance, upon illumination the CO2 concentration in the leaf intercellular air spaces is decreased by photosynthesis, resulting in decreased CO2 levels in the guard cells, which triggers stomatal opening. CO2 can then enter the leaf and photosynthesis can continue.
Light: The photoreceptors in stomatal cells can perceive certain light wavelengths, especially in the blue region of the visible spectrum. Activated by such receptors, stomata open with increasing light intensity. Stomatal conductance in the morning increases earlier than photosynthesis, which is only activated when the sun has risen further and with a larger proportion of red light. Therefore, stomatal opening caused by light does not adversely limit the flow of CO2 for photosynthesis during the day.
Temperature: Stomata close at low temperatures and open at higher temperatures. Increasing temperature can override the closing effects of increased CO2 levels and darkness. For example, increasing the temperature from 27oC to 36oC resulted in stomatal opening of Xanthum leaves in the dark without removal of CO2.
3. Stomatal resistance/conductance
We now turn to the resistance/conductance that each component of the leaf-atmosphere pathway offers to water vapor flow. Reference values are given in Table 1. The stomata are an obvious first choice for discussion. In the simplest case of a circular cylinder or other shape of circular cross-section, the resistance of the stomatal pore is given by 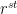
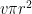


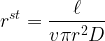
Because the pore area is much less than the leaf area, there is a zone close to the pore where the lines of flux converge on the pore. Therefore, this zone of the boundary layer is not effectively utilized and there is an extra resistance or ‘end effect’ associated with each pore. The magnitude of this extra resistance can be derived from three-dimensional diffusion theory to be proportional to the pore radius and ultimately equal to 

As an example, suppose we wish to determine the stomatal diffusion resistance and the corresponding conductance for water vapor flow on a leaf with 225 stomata/mm2 on each surface, each pore being 12 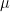

For each surface, 
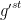
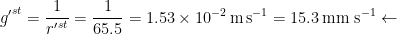
Table 1. Typical conductance values for components of the leaf-atmosphere pathway
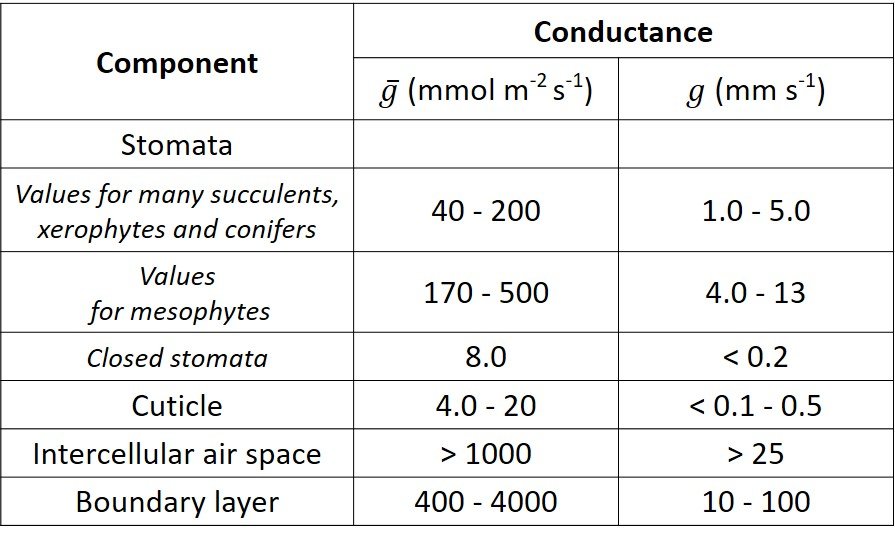
4. Other conductances/resistances
Some water molecules diffuse across the waxy cuticle of the epidermal cells; we can thus speak of cuticular transpiration, which involves both liquid water and water vapor. The cuticle is in parallel with the stomata, which is to say that water vapor can leave the leaf either by crossing the cuticle or by going through the stomatal pores. The conductance for cuticular transpiration, 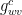

Another conductance to consider in the diffusion of water vapor in plant leaves is that of the intercellular air spaces, which usually account for about 30% of the leaf volume. Its value can be calculated with the ratio
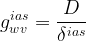
where D is the diffusion coefficient of water vapor in air and 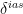
The fourth and last conductance/resistance component we must account for stems from the boundary layer, the thin stratum of disturbed airflow located between the surface of the leaf and the free-stream air. The thickness of the boundary layer is affected by the roughness of the leaf, the wind speed, and the distance along the leaf. Its contribution to resistance is relatively small. For a deeper insight on boundary layers in external flow, refer to one of our free materials on transport phenomena.
Three of the foregoing conductance components – stomatal, cuticular, and intercellular air space – together constitute the leaf conductance. Some values for leaves of select species are given in Figure 3.
Figure 3. Maximum leaf conductance in different groups of plants. The lines cover about 90% of individual values reported, and the open circles represent group average conductances.
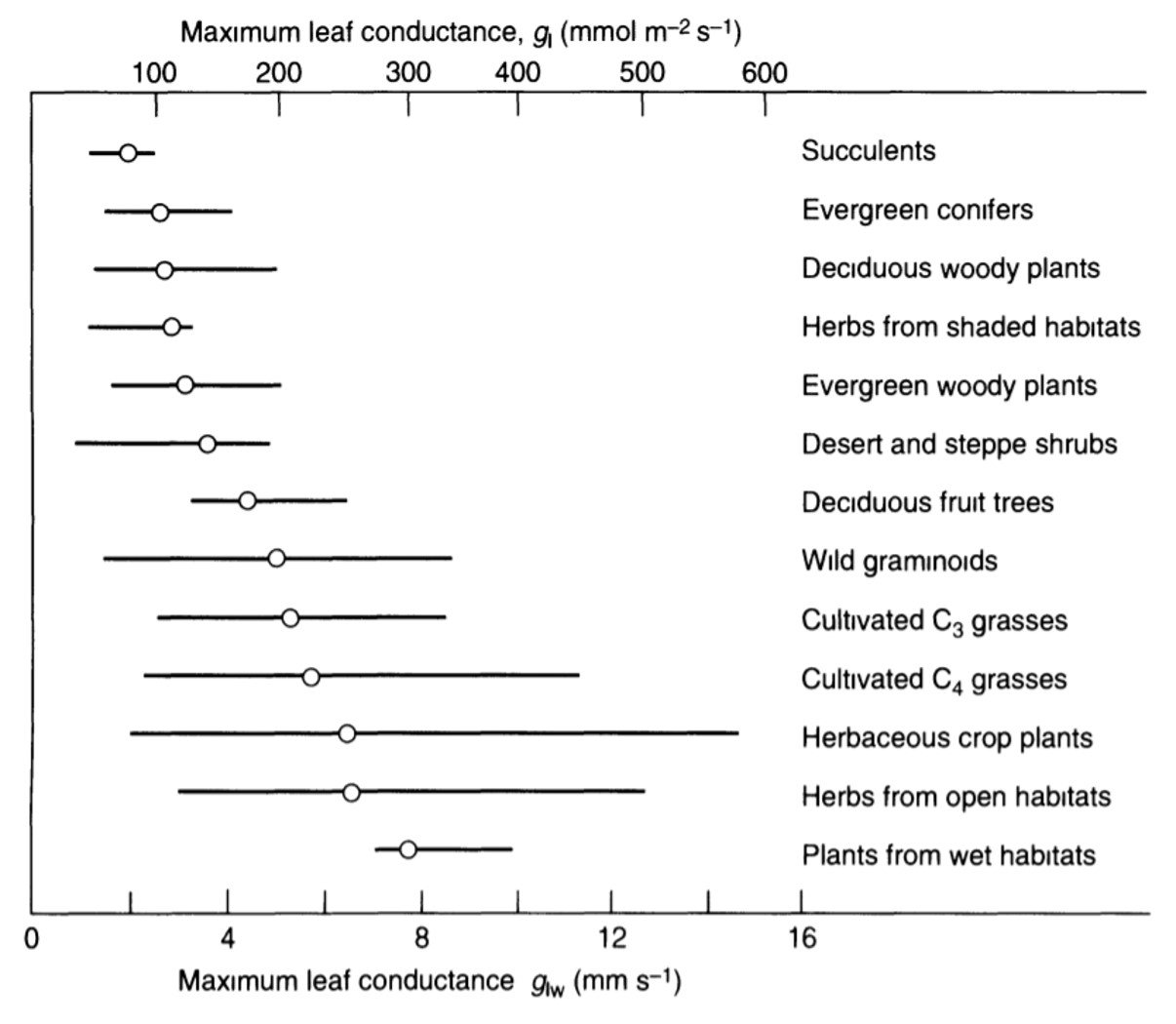
We have thus introduced the four conductance/resistance components involved in flow of water vapor across a leaf. As noted by Nobel (2009), two such components are strictly anatomical (intercellular air spaces and cuticle), one depends on anatomy and yet responds to metabolical as well as environmental factors (stomata), and one depends on leaf morphology and wind speed (boundary layer). We can assemble a hypothetical conductance pathway to represent flow of water vapor across the leaf, as shown in Figure 4.
Figure 4. Conductance arrangement for water vapor transpiration.
![]()
Let 

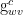
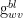


Next, we note that the leaf conductance 
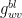
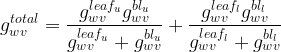
Having obtained the conductances, we can establish the net flux density 

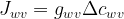
where 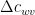
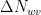
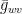

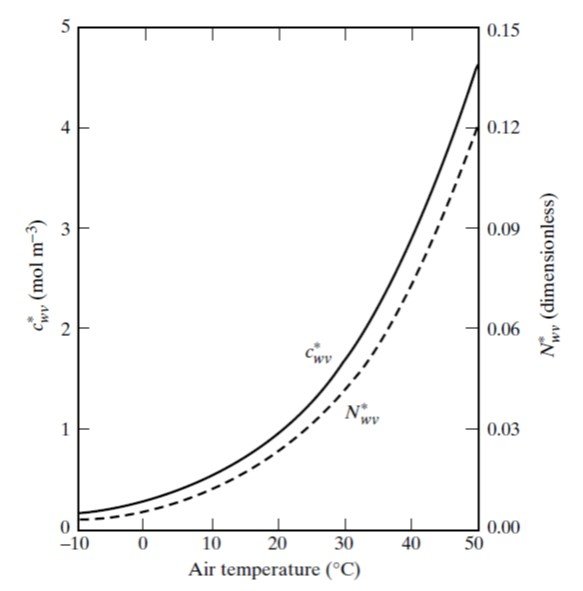
The mole fraction of water vapor can also be read from Figure 5.
Figure 5. Variation in saturation values of water vapor concentration (

Example (Modified from Nobel, 2009)
Suppose a leaf has the following conductances.
• Conductance of water vapor in the lower-surface boundary layer, 
• Conductance of water vapor in the lower-surface stomata, 
• Conductance of water vapor in cuticle,
• Conductance of water vapor in intercellular air spaces, 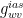
A) What is 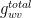
B) What would be the
C) What is
D) What is the total water vapor conductance if 30% of the diffusion flux is through the upper epidermis?
E) Suppose that the leaf temperature is 30oC, the air in the cell wall pores where the water evaporates is at 90% relative humidity, and the turbulent air water vapor concentration 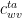
F) What are 
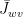

A) We begin by calculating the water vapor conductance of the leaf,
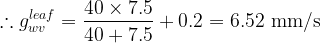
The total conductance of the leaf, observing that water vapor diffuses out only across the lower surface of the leaf, is then
B) If the cuticular pathway is ignored, the remaining three conductances are connected in series; that is,


If the intercellular air spaces are ignored, stomatal and cuticular conductances are in parallel (

and



If both cuticular and intercellular air spaces are ignored, we are left with a series connection of stomatal and boundary layer conductances, so that



C) The conductance of the upper surface here equals the conductance of the lower surface, determined in A, and is in parallel with it. In mathematical terms,

D) In this case, 70% of the diffusion flux stems from the lower epidermis. Using the total conductance calculated in part A, it follows that

E) The diffusion flux is given by
Conductance 

F) Appealing to the ideal gas law and noting that 1 m³ = 1000 L, we have

Referring to the dotted curve in Figure 5, we read a mole fraction 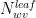


References
• JONES, H. (2014). Plants and Microclimate. 3rd edition. Cambridge: Cambridge University Press.
• LAMBERS, H., CHAPIN III, F., and PONS, T. (2008). Plant Physiological Ecology. 2nd edition. Berlin/Heidelberg: Springer.
• NOBEL, P. (2009). Physicochemical and Environmental Plant Physiology. 4th edition. London: Academic Press.
• WILLEY, N. (2016). Environmental Plant Physiology. New York: Garland Science.
• WILLMER, C. and FRICKER, M. (1996). Stomata. 2nd edition. Berlin/Heidelberg: Springer.

